
Teratologische Analyse eines missgebildeten Schafschädels aus der zoologischen Sammlung der Ruhr-Universität Bochum

(alle Fotos Copyright by Julian Lennartz, mit freundlicher Unterstützung der Ruhr-Universität Bochum und dem Leiter der zoologischen Sammlung Holger Bäcker)
1. Einleitung
Im Rahmen dieser Arbeit wird ein Tierschädel aus der zoologischen Sammlung der Ruhr-Universität Bochum vorgestellt. Das Objekt zeigt deutlich Erkennbare morphologische Veränderungen und Missbildungen, besonders im Bereich des Kiefers und der Augen. Im Rahmen dessen wird zunächst das biologische Fachgebiet der Teratologie kurz vorgestellt. Dies ist nötig um eine Begriffsdifferenzierung der verschiedenen Ausmaße von morphologischen Veränderungen vorzunehmen und verschiedene Faktoren, die zu Missbildungen führen können, kurz zu beleuchten.
Dann wird der Schädel anhand der makroskopischen Bestimmungsmerkmale eingeordnet und einer Spezies zugewiesen. Anhand einer ausführlichen fotografischen Dokumentation wird der Schädel beschrieben und mit dem Schädel eines gesunden Tieres der gleichen Spezies verglichen. Die fotografische Dokumentation ist im Anhang zu finden. Leider war es aufgrund verschiedener Umstände nicht möglich, den Schädel des gesunden Tieres während der Dokumentation auszumessen. Die Bestimmung und der Vergleich sollte aber dennoch gelingen.
Anhand der Auffälligkeiten am Schädel soll im nächsten Schritt eine Diagnose vorgenommen werden. Dies gestaltete sich aufgrund des fehlenden restlichen Körpers des Tieres als nicht immer ganz einfach. So kommen demnach drei verschiedene Syndrome in Frage, die sich alle ähneln, aber ohne den restlichen Körper des Tieres und genetische Analysen nicht differentialdiagnostisch voneinander unterschieden werden können.
Am Ende folgt eine Diskussion und Interpretation der Beobachtungen, ebenso wie eine Einschätzung ungeklärter Fragen, die entweder aufgrund fehlender Informationen oder wegen des Umfangs dieser Arbeit nicht weiter erläutert werden konnten.
2. Einführung in die Teratologie
Die Teratologie beschäftigt sich mit der Genese von Missbildungen durch Umweltfaktoren, seien diese endogen oder exogen. Bereits die Definition des Begriffs Missbildung gestaltet sich schwierig, da nicht klar zwischen dem „Normalen“ und „Abnormen“ getrennt werden kann (Rosenbauer 1969, S. 99). Den absoluten Archetypen einer Spezies gibt es nicht. Individuen unterscheiden sich untereinander. So sind bestimmte Abweichungen in der Erscheinung innerhalb einer Spezies, zum Beispiel in Größe, Proportionen und Gewicht, Stimmlage, Fell-, Feder-, Schuppen- oder Flügelfarbe häufig zu beobachten.
Höpker zitiert ein seinem Werk Christiansen und Smith und gibt folgende Begriffsdefinition an:
• „Malformation (Missbildung)
Ein primärer Strukturdefekt, der aus einem lokalisierten Fehler der Morphogenese resultiert (z.B: Lippenspalte).
• Deformation (Formänderung)
Eine Änderung in der Gestalt und/oder Struktur eines zunächst regelrecht angelegten Körperteiles (z.B. Torticollis).
• Anomalie
Eine Mißbildung zusammen mit den hieraus ableitbaren strukturellen Veränderungen (z.B. Robin'-Anomalie).
• Mißbildungssyndrome
Mißbildungsmuster, die überwiegend die gleiche Ätiologie aufweisen und meistens nicht als das Ergebnis eines einzelnen lokalisierten Fehlers in der Morphogenese erklärt werden können (z.B. Down'-Syndrom).
• Assoziation
Muster von Mißbildungen, welche noch nicht als Syndrom oder Anomalie angesehen werden können. Mit zunehmendem Wissen könnte eine Assoziation als ein Syndrom oder eine Anomalie eingeordnet werden (z.B. Hemihypertrophie mit Wilms'-Tumor)“
(Höpker 1984, S. 5-6).
Anhand dieser Unterteilung kann bei dem vorliegendem Objekt von einer Missbildung und einer Anomalie gesprochen werden. Zweiteres, weil sich aus den Ausprägungen der vorhandenen Missbildung sowohl strukturelle, als auch funktionelle Veränderungen ergeben. Wie sich im Verlauf dieser Arbeit herausstellen wird, scheinen sich alle in Frage kommenden Erkrankungen auf den Schädel zu beschränken.
Missbildungen werden als Kyematopathien bezeichnet. Damit sind alle nicht ausgeheilten vorgeburtlichen Erkrankungen gemeint. Abhängig vom Zeitpunkt ihres Auftretens wird nach Blastopathien, Embryopathien, Fetopathien und Placentopathien unterschieden (Rosenbauer 1969, S.101). Fehlbildungen die nach der Geburt auftreten fallen in die Postnatalpathologie und sind hier nicht Untersuchungsgegenstand. Ursachen für Missbildungen können endogener wie exogener Natur sein. Allerdings sind nur etwa 10% aller Missbildungen durch exogene und weitere 10% durch endogene Faktoren bedingt, wodurch die Ursachen für den weitaus größeren verbleibenden Teil von 80% einer komplizierten Wechselwirkung beider Ursachen unterliegen (Rosenbauer 1969, S. 102).
Die exogenen Ursachen werden von Rosenbauer in primär chemische, primär physikalische, primär iatrogene, primär durch Erkrankungen bedingte und unbekannte Faktoren unterschieden (Rosenbauer 1969, S. 103). Zu den physikalischen Faktoren gehören mechanische wie Nabelschnurumschlingungen oder Lageanomalien des Embryos, ionisierende Strahlen wie Röntgen- und Gammastrahlen, Ultraschall und Kurzwellen, thermische Faktoren und Sauerstoffmangel (Rosenbauer 1969, S. 107-113). Ein durch S. P. Hicks durchgeführtes Experiment zeigte, dass insbesondere Röntgenstrahlen, sowie Ultraschall und Kurzwellen bei Tieren, in diesem Fall bei Mäusen, teratogene Effekte auslösen (Rosenbauer 1969, S. 110). Bei Kaninchen und Ratten treten vor allem Extremitäten- und Schwanzmissbildungen auf, insofern die Durchflutung nach dem zehnten Trächtigkeitstag stattfindet; vorher sind eher Augenmissbildungen die Folge (Rosenbauer 1969, S. 111). Weiterhin reagieren Kaninchen im Experiment empfindlich auf Hyperthermien. Hyperthermien können sowohl exogene Ursachen, wie Bestrahlung, als auch endogene Ursachen, wie Fieber der Mutter während der Tragzeit haben (Rosenbauer 1969, S. 111). Sauerstoffmangel ist bei allen Wirbeltieren, den Menschen mit einbezogen, eine zu vielen unterschiedlichen Missbildungen führende Noxe (Rosenbauer 1969, S. 111ff.). Dieser führt vor allem zu Missbildungen des Herzens und der Wirbelsäule, aber auch des zentralen Nervensystems. (Rosenbauer 1969, S. 111-112). Sauerstoffmangel kann auch durch Herz- oder Gefäßmissbildungen der Mutter bedingt sein (Rosenbauer 1969, S. 118).
Zu den primär chemischen Faktoren zählen beim Menschen vor allem Medikamente, wobei es hierbei schwierig ist, diese von anderen chemisch induzierten Missbildungen abzugrenzen (Rosenbauer 1969, S. 113). Vor allem Trypanblau wurde tierexperimentell untersucht und erzeugt unter anderem Hydrozephalie, Spina bifida, Schwanzedefekte, Augendefekte und Ohrmissbildungen, Herzmissbildungen, Gaumenspalten, Meningozelen, Analatresien und weitere Fehlbildungen, womit Disazofarbstoffe, zu denen das Trypanblau, aber auch Afridolblau gehört, als teratogene Substanzen eingestuft werden können (Rosenbauer 1969, S. 114). Auch anorganische Salze wie Selen und Mangan können zu Missbildungen führen (Rosenbauer 1969, S. 114). Da seit dem Thalidomidvorfalll alle Medikamente für den Menschen zunächst an Tieren getestet werden, haben Medikamente wie Meclizin bei Tieren ebenfalls teratogene Effekte gezeigt (Rosenbauer 1969, S.118).
Viruserkrankungen wie Tollwut, Influenza und Miyagawanella psitacci führen bei Tieren ebenfalls zu Missbildungen (Rosenbauer 1969, S. 116). Verschiedene Mangeldiäten zeigen im Tierversuch eine ganze Reihe unterschiedlicher Fehlbildungen und so führt eine Vitamin-A-freie Kost bei Schweinen zu Mikrophthalmie oder Anophthalmie (Rosenbauer 1969, S.116). Der Einfluss von Stoffwechselstörungen der Mutter auf den Keim zeigt sich eher durch die Gabe der entsprechenden Medikation zur Behandlung der Störung und wird nicht weiter behandelt (Rosenbauer 1969, S.118).
3. Beschreibung des Objekts
3.1. Bestimmung der Spezies
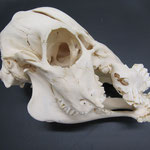
Das Tier gehört in jedem Fall zu den Wirbeltieren und den Mammalia. Das Objekt zeigt im Vergleich zum gesunden Tier einen deutlichen vergrößerten und fehlgestellten Zahnbesatz, sowohl im Ober- als auch Unterkiefer. Der Oberkiefer liegt ohne Schneidezähne vor und zeigt auch keine Hinweise auf verlorengegangene Schneidezähne, wodurch von einem Paarhufer (Artiodactyla) ausgegangen werden kann (Brohmer 2009, S. 639). Darauf lassen ebenfalls die großen Kauflächen der Backenzähne schließen. Vor den Augen liegt eine flache, rundliche Tränenbeingrube, was weitere Bestimmungsmerkmale unnötig macht und von einem Hausschaf (Ovis ammon) ausgegangen werden kann (Brohmer 2009, S. 653).
3.2. Makroskopische Beobachtung
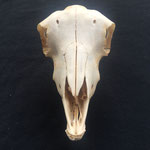
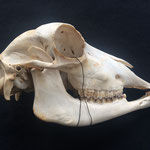
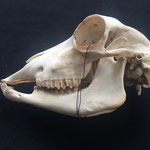
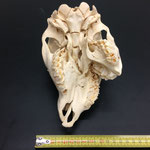
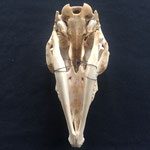
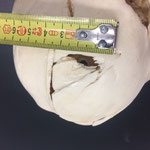
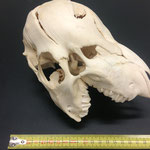
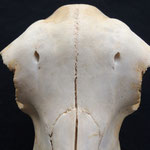
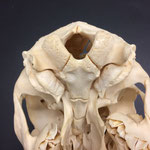
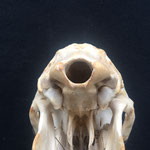
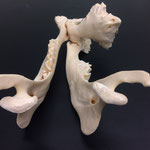
Makroskopisch zeigen sich deutliche Schädeldeformationen. Der Schädel hat ein zweites Paar Augen, sowie eine weitere Oberkieferstruktur ausgebildet. Vor allem der Hirnschädel weist Spalten und Löcher auf (Abb. 1). Das Nasenbein und die beiden Zwischenkieferbeine sind nach lateral links um etwa 30° verdreht. Lateral rechts entspringt etwa am Ende der rechten Backenzahnreihe eine weiterer Oberkiefer mit einer Gaumenlänge von etwa 7 cm, der jedoch kein Nasenbein ausgebildet hat (Abb. 2). Dieser Oberkiefer weist zwei weitere Zahnreihen auf, die jedoch zur Seite abstehen (Abb. 5). Die Ebene des Kiefers ist in Richtung des Gaumenbeins verdreht, sodass die Oberseite der beiden Oberkieferbeine nach lateral rechts zeigt (Abb. 5). Das Nasenbein und die Zwischenkieferbeine des eigentlichen Oberkiefers sind somit von dieser zusätzlichen Struktur abgewandt.
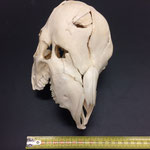

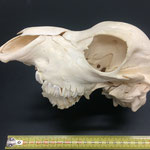

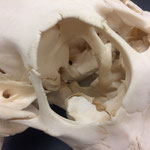


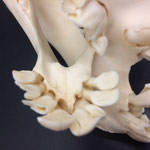



Auffallend ist ein eingewachsener, etwa 3 cm breiter Knochensplitter, der an der medialen Fontanelle anliegt und etwa im mittleren Stirnbereich vorzufinden ist (Abb. 6). Die rechten Ränder sind ausgefranst. Das Stück liegt etwas in den Hohlraum des Hirnschädels hineinversetzt. Dies lässt auf eine Kopfverletzung schließen. Am Hinterhaupt, etwa im oberen im Bereich des Zwischenscheitelbeins ist eine weitere Öffnung zu sehen (Abb. 7 u. 7B). An dieser Stelle laufen die offenen Fontanellen zusammen, welche wahrscheinlich durch Knorpel verschlossen waren, der nach dem Tod des Tieres verrottete.
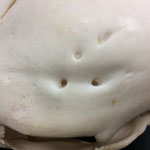
Das Tier zeigt Anlagen für vier Augenhöhlen. Das linke Auge ist normal ausgebildet. An der Position, an der normalerweise das rechte Auge liegen würde, lassen sich zwei weitere Augenhöhlen identifizieren, die linke davon etwas kleiner als die rechte. Die gesamte Struktur ist etwa 4 cm breit (Abb. 8). Erst dann folgt eine weitere große Augenhöhle, die etwa mittig am oberen Ende der Naht der Oberkieferbeine des zweiten Oberkiefers anliegt (Abb. 2 u. 8). Es liegt nahe, dass das linke große Auge, auch eben dieses des Hauptgesichtes war, die linke kleine Augenhöhle eigentlich die rechte des Hauptgesichtes sein müsste, die rechte kleine eigentlich die linke des zweiten Gesichtes und die rechte große eben diese des Nebengesichtes war. Es wäre, wie sich später noch zeigen wird, ungewöhnlich, wenn das zweite Gesicht zwischen den beiden Augen und dann auch noch medial verschoben mitten aus dem ersten hinauswachsen würde. Innerhalb des Gesichtsschädels sind die drei rechten Augenhöhlen nur unzureichend voneinander abgetrennt und weisen deutliche Zugänge zum Hirnschädel auf, die von den normalen Ausmaßen eines Kanals für den Sehnerv abweichen (Abb. 9). Betrachtet man die Anlage der Augen und derStirnbeine genauer, wirkt es so, als wäre lateral rechts ein Schädelknochen zu viel angelegt worden, wodurch eine Fontanelle ebenfalls zu viel ist (Abb. 2 u. 7).
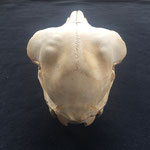
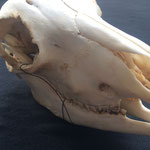
Die Durchtrittsöffnungen für Nerven und Arterien, die normalerweise im Stirnbein angelegt sind, fehlen auf der rechten Seite. Auf der linken Seite, lassen sich dagegen vier Öffnungen erkennen (Abb. 10). Das untere Hinterhauptbein ist in drei Teile zerbrochen oder niemals als zusammenhängende Struktur angelegt gewesen (Abb.4). Das Hinterhauptloch ist regulär geformt, allerdings zeigen die Hinterhauptsgelenkhöcker deutliche Abnutzungserscheinungen (Abb 11 u. 11B). Ebenso ist der Schädel im Bereich um das Hinterhauptloch deutlich breiter angelegt, als beim normalen Tier (Abb. 11 u. 11B). Die äußeren Gehörgänge sind leicht nach hinten, statt im 90°-Winkel zur Sagittalebene gerichtet (Abb. 3 u. 3B).
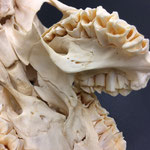
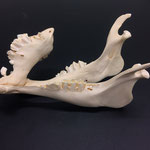
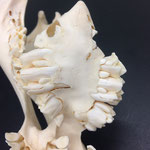
Auch am Unterkiefer hat sich direkt hinter den Schneidezähnen auf der rechten Seite eine weitere Unterkieferstruktur ausgebildet (Abb. 12, 13 u. 14). Dieser ragt in einem 45° Winkel nach oben. Die beiden Knochenteile haben sich in der Entwicklung nicht gespalten, sondern bilden eine feste zusammenhängende Struktur, die mit zwei Reihen Backenzähnen besetzt sind, die ebenso wie bei der zusätzlichen Oberkieferstruktur von der Mittellinie hinweg ragen (Abb. 15). Die Struktur entspringt zwischen dem normalen und einem zusätzlichen Schneidezahnbesatz (Abb. 16). Die Gelenkansätze am Unterkiefer sind asymmetrisch ausgebildet (Abb. 14). Der gesamte rechte Unterkieferknochen ist nach rechts aussen verdreht (Abb. 14).
Die Durchtrittsöffnungen für Nerven und Blutgefäße sind im eigentlichen Unterkiefer regulär angelegt, bei der zusätzlichen Struktur finden sie sich auf der Rückseite entlang der Mittellinie etwa mittig auf beiden Seiten und am oberen Ende der Struktur (Abb. 12, 13 u. 14). Der gesamte Zahnbesatz des Tieres weist nur sehr geringe, bis gar keine Abnutzungserscheinungen auf. Die Zähne wirken porös.
Im Gesamthabitus lassen sich die Gelenke von Ober- und Unterkiefer nicht richtig miteinander verbinden (Abb. 17). Ein vollständiger Kieferschluss ist nicht möglich (Abb. 17). Lediglich die linken Backenzähne von Ober- und Unterkiefer liegen lose aufeinander auf. Die Schneidezähne des Unterkiefers schließen nicht mit den Zwischenkieferbeinen ab, sondern ragen in Form eines Überbisses über diese hinaus (Abb. 18). Auffallend ist, dass die zusätzliche Unterkieferstruktur genau in die Lücke zwischen den beiden Oberkieferstrukturen ragt. Lateral rechts ragt der zweite Oberkiefer über den Unterkiefer hinaus (Abb. 19). Auch hier ist kein Kieferschluss erkennbar. Die Backenzähne des Unterkiefers beginnen erst nach denen des normalen Oberkiefers (Abb. 19). Die Zähne des zweiten Oberkiefers haben mit denen des Unterkiefers gar keinen Kontakt. Die Zähne des zweiten Unterkiefers haben mit keiner anderen Struktur Kontakt.
3.3. Abmessungen
Die Breite des Schädels beträgt etwa 14 cm, gemessen vom linken Schuppenbein bis zum Oberkieferbein des zweiten Oberkiefers, der über das rechte Schuppenbein hinausragt (Abb. 1 u. 4). Die Condylobasal-Länge beträgt etwa 19 cm (Abb. 2 u. 3). In beiden Oberkiefer beträgt die Länge des Backenzahnbesatzes etwa 7cm.
4. Diagnose
Aufgrund der zusätzlichen Kieferstrukturen, die in anderen Entwicklungsstadien als die normalen Kieferstrukturen vorliegen, kann von einer asymmetrischen zusammenhängenden Doppelbildung ausgegangen werden (Stünzi 1985, S. 634). Diese entstehen, wenn die Blastozyste beginnt sich zu spalten, aber diese Spaltung unvollständig bleibt. Bei symmetrischen Doppelbildungen können zwei vollständige Körperachsen entstehen, was eineiige Zwillinge zur Folge hat. Trennen sich die Körperachsen nicht vollständig voneinander, entstehen siamesische Zwillinge (Stünzi 1985, S. 634). Somit kommt für das vorhandene Objekt ein Diprosopus, eine siamesische Zwillingsform und eventuell ein parasitärer Zwilling in Frage. Fraglich bleibt, ob es sich um eine Blastopathie oder eine Embryopathie handelt. Die Klärung dieser Frage würde die Diagnosestellung deutlich vereinfachen. Der Diprosopus ist, ausgehend von den etwas widersprüchlichen und teilweise auch ungenauen Angaben der verwendeten Literatur eher eine Embryopathie, während die Zwillingsformen den Blastopathien zuzuordnen sind.
4.1. Diprosopus
Da nur der Schädel des Tiers vorliegt und somit keine anderen Symptome der Missbildung am Körper des Tieres festgemacht werden können, ist die wahrscheinlichste Diagnose eine craniofaziale Vervielfältigung, auch Doppelgesicht oder Diprosopus genannt und gehört zu den symmetrischen Doppelfehlbildungen. Diese betrifft, je nach Ausprägung, ausschließlich das Gesicht, beziehungsweise den Schädel und in seltenen Fällen auch die Halswirbelsäule (Hu/Helms 1999, S. 4873, Stünzi 1985, S. 636). Katzen mit diesem Syndrom werden auch Januskatzen genannt (Abb. I). Diese seltene Fehlbildung und auch Cyklopien werden durch das Sonic-Hedgehog-Protein (SHH) hervorgerufen (Hu/Helms 1999, S. 4873). Die Tiere haben einen ansonsten normal ausgebildeten Körperbau, allerdings können Neuralkanaldefekte, Fehlbildungen des Herzens und Anencephalie auftreten (DeRuiter 2011). Ist das Gehirn ausgebildet zeigen sich oft duplizierte Bereiche (DeRuiter 2011). Die Fehlbildung kann in verschiedenen Ausprägungen auftreten, von einem Gesicht mit mehreren Augenpaaren, über komplette Anlagen zweier Kiefer, wie bei der Januskatze in Abb. I, bis hin zu zwei vollständig ausgeprägten, voneinander getrennten Köpfen (Hu/Helms 1999, S. 4873). Stünzi weist aber, im Gegensatz zu den anderen Autoren, darauf hin, dass die Gesichtsteile sich auch asymmetrisch entwickeln können und zusätzliche Kieferstrukturen nicht vollständig sein müssen, während der Hirnschädel einfach angelegt ist, wobei der Diprosopus dennoch zu den symmetrischen Doppelfehlbildungen gezählt wird (Stünzi 1985, S. 636). Diese Beschreibung passt zu dem vorliegenden Objekt, da hier eine asymmetrische Vervielfältigung von Gesichtsteilen vorliegt und sich die Kopie auch nicht entlang der Medialebene entwickelt hat. Die Entstehung eines Diprosopus wird häufig mit der eines siamesischen Zwillings in Verbindung gebracht, vor allem dann, wenn sich die beiden Gesichter oder Köpfe sehr ähnlich sind (DeRuiter 2011). Dabei entsteht die Teilungslinie in der Medialebene, sodass in der Embryonalentwicklung zunächst der Kopf und dann der Körper geteilt wird, diese Teilung aber unvollständig bleibt und so die beiden Gesichter entstehen (DeRuiter 2011). Allerdings haben nur 0,4% aller menschlichen siamesischen Zwillinge einen Diprosopus (DeRuiter 2011). Daten über die Häufigkeit bei Tieren liegen in der verwendeten Literatur nicht vor. Auch DeRuiter weist auf das SHH-Protein hin. In Tierversuchen bildeten Hühner einen Diprosopus aus, wenn sie während ihrer Entwicklung im Ei einer erhöhten Dosis SHH-Protein ausgesetzt wurden, wohingegen eine Cyklopie auftrat, wenn zu wenig SHH im Organismus vorhanden war (DeRuiter 2011, Hu/Helms 1999, S. 4874). SHH beeinflusst während der Embryonalentwicklung die morphologische Gestaltung des Schädels, zunächst im axilaren Mesondoderm und später im Bereich des Nasenbeins und des Oberkieferbeins, vor allem aber der mediolateralen Breite, die, wenn zu groß zu Diprosopus und, wenn zu klein, zu Cyklopie führt (Hu/Helms 1999, S. 4874, 4880). SHH nimmt direkten Einfluss auf das Ektoderm, dass für die Gesichtsgestaltung zuständig ist, wobei unklar ist, in welcher Form das Ektoderm die Gesichtsentwicklung bestimmt (Hu/Helms 1999, S. 4875). Der Einfluss von SHH beschränkt sich lediglich auf das Ektoderm der Frontonasal- und der Maxillarentwicklung und insbesondere auf die Entwicklung des Gaumenbeins (Hu/Helms 1999, S. 4876). Im Tierversuch zeigte sich jedoch auch, dass der Einfluss von SHH auf das Ektoderm abhängig vom Entwicklungsstadium der Tiere ist. Wird das Ektoderm in frühen Entwicklungsstadien chirurgisch entfernt, heilt es innerhalb kurzer Zeit wieder ab und die Gesichtsentwicklung verläuft normal. In späteren Entwicklungsstadien führt die Entfernung jedoch zu morphologischen Veränderungen des Gesichts, was ebenfalls zeigt, dass das Ektoderm selbst das SHH-Protein bildet (Hu/Helms 1999, S. 4876). Warum jüngere Embryos dazu fähig sind, SHH-bildendes Gewebe nachzubilden, ältere jedoch nicht, bleibt unklar (Hu/Helms 1999, S. 4877). Fest steht damit jedoch, dass der Einfluss von SHH-bildendem Gewebe auf die craniofacielle Entwicklung erst in den späteren Stadien der Embryonalentwicklung wirksam wird. Pharmakologische Dosen von Retinoiden und Cholesterinanaloga beeinflussen die Signalwirkung von SHH, was ein Beweis für den Einfluss des Proteins auf die craniofacielle Entwicklung ist (Hu/Helms 1999, S. 4880). Pflanzliche Alkaloide beeinflussen vor allem bei Schafen die Gesichtsentwicklung, da sie die Signalwirkung von SHH stören und es so zu Lippen-Kiefer-Gaumenspalten kommen kann (Hu/Helms 1999. S. 4880). Dies zeigt ebenfalls, dass ein Diprosopus unabhängig von der Entwicklung siamesischer Zwillinge auftreten kann. Allerdings weisen Hu und Helms auch darauf hin, dass nicht klar gesagt werden kann, ob eine Vergrößerung der mediolateralen Breite allein, ob durch SHH-Einfluss oder nicht, tatsächlich der Ursprung für einen Diprosopus ist oder dazu führen kann (Hu/Helms 1999, S. 4881).
Da nicht genau klar ist in welcher Phase der Embryonalentwicklung der Einfluss von SHH einen Diprosopus auslöst, muss davon ausgegangen werden, dass verschiedene Teratogene und Entwicklungsstörungen zu diesem seltenen Syndrom führen. (DeRuiter 2011).
Da der hier untersuchte Schädel durch die Fehlbildungen etwas größer als ein normaler Schafschädel ist, kann davon ausgegangen werden, dass das Tier zum Zeitpunkt des Todes ausgewachsen und somit lebensfähig war. Die in Abb. I gezeigte Katze wurde im Jahre 2011 zwölf Jahre alt (http://www.spiegel.de/panorama/gesellschaft/kurz-krass-katze-mit-zwei-gesichtern-feiert-geburtstag-a-789270-2.html). Dies zeigt, dass Tiere mit dieser Missbildung durchaus lebensfähig sind, insofern keine lebenswichtigen Organe betroffen sind.
4.2. Siamesische und parasitäre Zwillinge
Spaltet sich die Blastozyste in der Entwicklung nicht vollständig und symmetrisch auf, spricht man von einer inaequalen unvollständigen Spaltung, oder einer asymmetrischen Doppelbildung (Stünz 1985, S. 638). Dabei kann ein Teil der Blastozyste einen vollständigen Organismus entwickeln, in dem sich der unvollständige zweite Organismus einlagert. Der Vollständig entwickelte Körper wird als Autosit, der unvollständige als Parasit bezeichnet und dieser kann sich an Kopf, Brust und Bauch anlagern (Stünzi 1985, S. 638). So kann es dazu kommen, dass sich die unvollständigen Teile des Parasiten in die Kieferknochen einlagern und so eine Verdoppelung der Kieferknochen (Dignathie) zu beobachten ist (Stünzi 1985, S. 638). Der parasitische Kieferteil wird als Epignathus bezeichnet und kann sich sowohl bindegewebsartig, als auch knöchern in den Schädel des Autositen einlagern (Stünzi 1985, S. 638). Da bei dem vorliegenden Objekt aber auch vor allem die Augenhöhlen das entscheidende Merkmal sind, trifft wohl eher ein Otognathus zu, bei dem sich rudimentäre Maul- und Augenteile im Bereich der Schläfe entwickeln; dies käme vor allem bei Rindern und Schafen vor (Stünzi 1985, S.638).
4.3. Mögliche Ursachen für die Missbildung
Die Faktoren, welche die Missbildung begünstigt oder ausgelöst haben sind aus der Literatur nicht klar zu benennen. Allerdings sind aus den Ausführungen von Hu und Helms in erster Linie endogene Faktoren naheliegend. Der Einfluss von SHH auf das Ektoderm ist zwar bewiesen, wie die Exposition jedoch ausgelöst wird, geht aus der Literatur nicht hervor. Genetische Ursachen kommen als endogene Faktoren ebenso in Frage wie Inzest. Als exogene Faktoren kommen Strahlung und primär chemische Expositionen in Frage, aber auch die Exposition mit pflanzlichen Alkaloiden.
5. Fehlereinschätzung und Fazit
Es konnte gezeigt werden, dass bei dem vorhanden Schafschädel eine deutlich erkennbare Anomalie vorliegt, deren Ursachen wahrscheinlich einer komplizierten Mischung aus endogenen und exogenen Faktoren unterlagen. Aufgrund der Ausprägung der Missbildung waren die wahrscheinlichsten Diagnosen der Diprosopus, eine Form eines siamesischen Zwillings, wobei hierfür der restliche Körper untersucht werden müsste, oder ein parasitärer Zwilling. Ob es zu Mischformen zwischen diesen Erkrankungen kommen kann ging aus der Literatur nicht hervor.
Anhand der Beschreibungen des Syndroms des Diprosopus und dem Vergleich mit anderen Tieren, die ähnliche Missbildungen ausgebildet haben, ist die Diagnose des Diprosopus sehr wahrscheinlich aber dennoch schwer zu stellen. Normalerweise liegt hierbei eine mediolaterale Duplikation vor. Das Gesicht des Schafes müsste also entlang der Medialeben kopiert worden sein. Demnach müsste ein zweites Nasenbein und eine vollständigere Anlage des Unterkiefers vorliegen. Allerdings wachsen die zusätzlichen Kieferstrukturen nicht entlang der Mediallinie. Der Diprosopus könnte in diesem Fall sehr schwach ausgebildet sein, so dass noch keine vollständige Kopie des Gesichts angelegt wurde. Allerdings lässt sich aus der Literatur erkennen, dass sich beide Gesichter zeitgleich entwickeln würden, was hier eindeutig nicht vorliegt. Speziell die zweite Oberkieferstruktur liegt in ihrer Entwicklung deutlich hinterher. Gleiches gilt für einen siamesischen Zwilling. Auch hier müssten beide Gesichter den gleichen Entwicklungszustand aufzeigen. Gerade dieser Entwicklungsdefizit macht eine Einordnung der Missbildung so schwierig. Bei parasitären Zwillingen lässt sich der Parasit gegenüber dem Autositen klar abgrenzen und liegt häufig in einer Art Gewebesack vor. In dem vorliegenden Fall hätte der Parasit dann aber mitten aus dem Gesicht des Autositen herauswachsen müssen. Der parasitäre Zwilling ist zusätzlich eher im Bereich des Abdomens zu finden. BBC berichtete über einen Fall eines Jungen, dem Teile eines parasitären Zwillings aus dem Hirnschädel entfernt wurden, doch auch hier lag das Gewebe des Zwillings klar abgegrenzt vor (http://news.bbc.co.uk/2/hi/health/7791321.stm). Sowohl ein Diprosopus, als auch ein parasitärer Zwilling ist wahrscheinlich. Beides erlaubt eine asymmetrische Anlage von zusätzlichen Gesichtsteilen, jedoch aufgrund unterschiedlicher Ursachen. Fraglich ist, ob hier nicht sogar ein bisher undokumentierter Einzelfall eines bisher nicht beschriebenen Syndroms vorliegt.
Zusätzlich ergeben sich weitere Unklarheiten. Das Geschlecht des Tieres lässt sich anhand des Schädels nicht bestimmen, ist für die Beurteilung der Missbildung jedoch irrelevant. Es ist anzunehmen, dass nur eine Zunge ausgebildet war. Da nur einer der beiden Unterkiefer aus zwei getrennten Knochen besteht, gibt es auch nur einen Ansatz für einen Zungengrund.
Unklar bleibt, ob die beiden mittleren Augenhöhlen mit Augäpfeln besetzt und ob diese sehfähig waren. Kanäle für die Sehnerven bis zum Gehirn sind jedenfalls vorhanden. Die asymmetrische Anordnung der beiden „großen“ Augen könnte die Stereoskopie des Tieres eingeschränkt haben. Vielleicht ist dadurch auch die „Kopfverletzung“ entstanden, indem das Tier mit dem Kopf irgendwo angestoßen ist. Der Angriff eines Artgenossen könnte ebenfalls dazu geführt haben. Ob das Tier von seinen Artgenossen aufgenommen oder verstoßen wurde, bleibt eine interessante, jedoch offene Frage. Ebenso, inwiefern die leibliche Mutter das Tier umsorgt hat.
Aufgrund der geringen Abnutzungserscheinungen des Zahnbesatzes, liegt die Vermutung nahe, dass das Tier auf andere Art ernährt wurde, als die natürliche Nahrungspräferenz bei Schafen. Da das Tier zum Zeitpunkt des Todes ausgewachsen war, aber wahrscheinlich keine feste Nahrung aufnehmen konnte, da der unvollständige Kieferschluss keine Kaubewegungen oder Abrupfen von Gräsern zulässt, kann angenommen werden, dass das Tier unter menschlicher Pflege aufgewachsen ist. Aufgrund des auffällig unvollständigen Kieferschlusses ist fraglich, ob das Maul durch die Lippen oder weitere Hautfalten verschlossen werden konnte. Falls dies nicht der Fall war, muss das Tier unter einer ständigen Mundtrockenheit gelitten haben. Es bleibt weiterhin unklar, ob das Tier durch die Missbildung Schmerzen leiden musste und wenn ja, ob diese durch menschliche Intervention gezielt gestillt wurden.
Literatur
Brohmer, P. (Begr.) (2009). Brohmer - Fauna von Deutschland. Ein Bestimmungsbuch unserer Heimischen Tierwelt (23. Auflage; Neu herausgegeben von M. Schaefer, 2009). Quelle & Meyer-Verlag
DeRuiter, Corinne
(2011). Diprosopus
(Craniofacial Duplication).Embryo Project Encyclopedia(2011-08-16). ISSN: 1940-5030 http://embryo.asu.edu/handle/10776/2286
Höpker, W. W. (1984). Mißbildungen, Interrelationen, Assoziationen und diagnostische Validität.Berlin: Springer Verlag
Hu, Diane / Helms, Jull A. (1999). The role of Sonic hedgehog in normal and abnormal craniofacial morphogenesis. San Francisco, USA: Department of Orthopaedic Surgery, School of Medicine, U-453, University of California at San Francisco, Printed in Great Britain © The Company of Biologists Limited
Rosenbauer, Karlheinz A. (Hrg) (1969). Entwicklung, Wachstum, Missbildungen und Altern bei Mensch und Tier. Stuttgart:Wissenschaftliche Verlagsgesellschaft mbH
Stünzi, H. (1985) Handbuch der speziellen pathologischen Anatomie der Haustiere – Band IV, Genitalorgane, Haut, Skelett (Vollständig überarbeitete Neuauflage) Berlin Verlag Paul Parey